
| Surirella biseriata (600-facher und 700-facher Zeitraffer) | Nahezu kreisförmige Bahn (150-facher Zeitraffer) |
Krümmung der Trajektorien am Beispiel Surirella biseriata
Im April 2017 isolierte ich eine Diatomee der Gattung Surirella in dem Flüsschen Lein (48°53'04.6"N 9°38'29.2"E), dem Abfluss des Aichstruter Stausees.
 |
Das Bild links zeigt die Diatomee in Valvenansicht. Ich ordne sie der Spezies Surirella biseriata zu (siehe http://cyclot.sakura.ne.jp/keisougazou/surirel/Surirell/sursei.html). Das Video links oben zeigt ein Beispiel der Bewegung der Diatomeen in Kultur in 600-fachem Zeitraffer. Diese Diatomeen bewegen sich meist sehr langsam. In Petrischalen aus Polystyrol lagen die schnellsten Bewegungen bei ca. 2,8 µm/s. Für ihre eigene Länge, die bei meinen Kulturen typisch etwas über 210 µm betrug, benötigten sie mehr als eine Minute. Bei Beobachtungen an senkrechten Glasflächen traten deutlich höhere Geschwindigkeiten auf.
Wie das Titelvideo links oben zeigt, bewegen sich die Surirella biseriata häufig nur ein wenig hin und her. Bereits in mäßig dicht besiedelten Kulturen bildet sich ein merklicher EPS-Film auf dem Substrat. Diatomeen gleiten darauf bei der geringsten Bewegung der Petrischalen und ihre aktive Bewegungsfähigkeit ist eingeschränkt. Zur Beobachtung der Bewegung empfiehlt es sich daher, ausreichend viele Diatomeen in eine frische Petrischale zu pipettieren und dort zu beobachten. Nach meinem Eindruck werden außerdem längere Bahnen vor allem von Diatomeen zurückgelegt, die noch nicht viele Reservestoffe eingelagert haben. Eine hohe Lichtintensität scheint für längere Bahnabschnitte ohne Richtungsumkehr ebenfalls nicht günstig zu sein.
Wenn die Diatomeen zwischen Umkehrpunkten nicht nur sehr kurze Strecken zurücklegen, ist unverkennbar, dass die Trajektorie gekrümmt ist. Offenbar befindet sich zwischen den Umkehrpunkten immer eine Raphen-Hälfte im Kontakt mit dem ebenen Substrat. Da die Krümmung der Raphe auf jeder Seite der Diatomee einheitlich ist, lässt sich die Krümmung der Trajektorie eindeutig einem der beiden Raphen-Abschnitte zwischen den Apices zuordnen. Oben rechts ist ein Beispiel zu sehen, bei dem sich die Diatomee ungefähr auf einer Kreisbahn bewegt. Zur Verdeutlichung ist ein Kreis eingeblendet. Den Abschluss des Videos bildet eine Überlagerung aller Frames. Offenbar wird die Krümmung der Bahn durch den außen liegenden Teil der Raphe bestimmt.
Man kann Surirella biseriata in Valvenlage (eine Valve liegt oben) und Gürtelbandlage (Gürtelband liegt oben) antreffen. Wenn die Diatomeen sich um die Apikalachse drehen, was oft zu beobachten ist, wechseln sie häufig zwischen diesen Lagen. Auch Zusammenstöße zwischen Diatomeen können einen Wechsel der Lage verursachen. Es soll nur die Bewegung in Valvenlage betrachtet werden, da hierbei gut beobachtbare und beschreibbare Bahnen durchlaufen werden. Bemerkenswert sind insbesondere die Krümmungen an den Umkehrpunkten.
Die Voraussetzungen für eine Analyse der Bahnen wie sie im Beitrag „Analyse der Trajektorien I“ vorgestellt wurde, sind wegen der außen liegenden stark gebogenen Raphen nicht gegeben. Diese Geometrie legt jedoch eine einfache Interpretation der Bewegungsmuster nahe.
Umkehr ohne Wechsel der Krümmungsrichtung
Hierbei bewegt sich eine Diatomee, die in Aufsicht eine Bahn im Uhrzeigersinn beschreibt, nach der Umkehr weiterhin im Uhrzeigersinn. Entsprechendes gilt für den Gegenuhrzeigersinn.
In der Skizze unten links sind zwei solche Umkehrpunkte hintereinander dargestellt. Die in den Bahnabschnitten aktive Raphe ist rot markiert. Sie steht mit dem Substrat in mechanischem Kontakt.
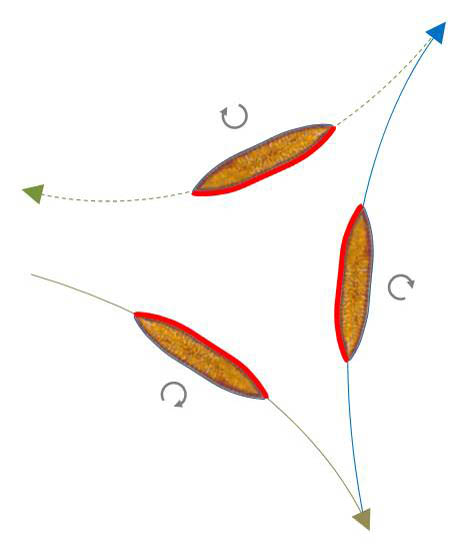 |
Über die Aktivität des anderen Raphen-Abschnitts lässt sich aus der Bewegungsbahn nichts aussagen. Offenbar wechselt bei der Umkehr die Seite, die sich in Kontakt mit dem Substrat befindet (Kippen um die Apikalachse). Zudem ist die Richtung der Aktivität der Raphen-Abschnitte unterschiedlich, so dass es zur Bewegungsumkehr kommt. Rechts daneben ist ein Video in 300-fachem Zeitraffer zu sehen, bei der eine solche Umkehr zu erkennen ist.
 |
Am Ende des kurzen Videos wird ein Bild eingeblendet, das aus Überlagerung aller Frames des Videos entstanden ist und die ganze Trajektorie zeigt.
Diese Form der Umkehr mit Erhalt des Drehsinns ist sehr häufig. Das folgende Bild (zum Vergrößern anklicken) zeigt eine Überlagerung der Frames eines Videos der Bewegung in einer Kultur. Man erkennt an vielen Stellen die charakteristischen Muster dieses Verhaltens.
Daraus folgt, dass sehr oft die Bewegung in den Raphen-Hälften gegensinnig ist. Die Aktivität des gesamten Raphensystems erfolgt in einem Drehsinn, gewissermaßen im „Kreis“.
Umkehr mit Wechsel der Krümmungsrichtung
Diese Form des Umkehrens wurde als typische Form bei Navicula vorgestellt. Es genügt bei Surirella biseriata, dass sich bei bestehendem Kontakt zwischen Raphe und Substrat die Richtung der Aktivität der Raphe umkehrt. Die Skizze unten links verdeutlicht wieder den Ablauf.
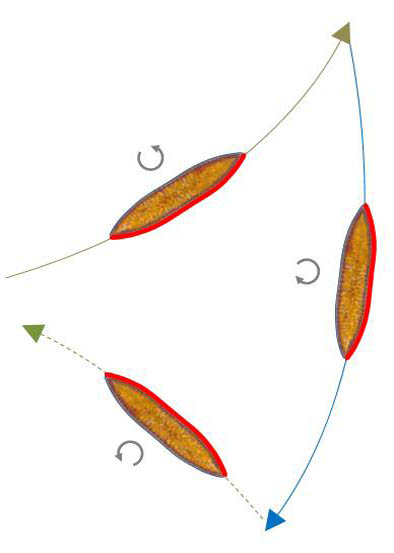 |
Im Video rechts daneben (150-facher Zeitraffer) ist zunächst eine Umkehr ohne Wechsel der Drehrichtung zu sehen, wie sie bereits vorgestellt wurde. Danach erfolgt der Wechsel der Richtung der Aktivität. Am Ende des Videos wird wieder das Überlagerungsbild eingeblendet. Ich konnte diese Art des Richtungswechsels nur gelegentlich beobachten. Dies kann auch daran liegen, dass solche Richtungswechsel in kurzer Folge geschehen, so dass die Bahnkrümmung zwischen den Umkehrpunkten nicht erkennbar ist.
Sigmoide Bewegungsbahnen
Bei Round et. al (2007) heißt es (S. 105): „Movement is directional, the path taken correspondingly fairly closely to the course of the raphe system (..) – curved where the raphe is curved (e.g. some Nitzschia species with eccentric raphe systems), straight where the raphe is straight (e.g. Navicula, Pinnularia), and even sigmoid where the raphe is sigmoid (e.g. Pleurosigma angulatum).”
Pleurosigma und Gyrosigma besitzen ein Raphen-System auf jeder Valve, das eine sigmoide Struktur mit dem Wendepunkt in der Mitte der Valve bildet. Eine sigmoide Bahn wird durchlaufen, wenn das Zentrum des Antriebs von einem Abschnitt mit positiver Krümmung zu einem mit negativer Krümmung wechselt und dabei die Fortbewegungsrichtung erhalten bleibt. Entsprechendes gilt für den umgekehrten Wechsel von negativer Krümmung zu positiver Krümmung. Offenbar ist dafür nicht die Anordnung der Raphenäste von Bedeutung, sondern das Vorkommen positiver und negativer Krümmungen. Ein sigmoides Raphen-System ist also nicht zwingend erforderlich. Deshalb kann man das Vorkommen sigmoider Trajektorien auch bei Surirella biseriata vermuten. Es genügt ein Wechsel des mechanischen Kontakts zum Substrat von einer Seite auf die andere, sofern die beiden Raphensysteme in derselben Richtung arbeiten, wie es die Skizze unten links zeigt. Die Trajektorie besitzt deshalb einen Wendepunkt am Ort des Wechsels des Kontakts.
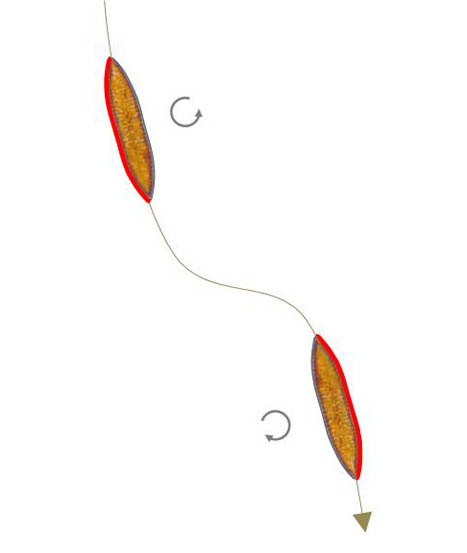 |
Im Video daneben (900-facher Zeitraffer) ist eine solche Bahn mit anschließendem Überlagerungsbild zu sehen.
Die oben beschriebene Umkehr der Bewegungsrichtung ohne Wechsel der Krümmungsrichtung der Trajektorie (Erhalt des Drehsinns) und das Auftreten von Wendepunkten beruhen auf dem Wechsel des mechanischen Kontakts zwischen Substrat von einer Raphenhälfte auf die andere. Das Umkehren mit Erhalt des Drehsinns ist wesentlich häufiger als das Auftreten von Wendepunkten. Man kann daraus schließen, dass die Aktivität der Raphen-Abschnitte an einer Valve nach dem Kippen meist entgegengesetzt und nicht parallel verläuft. Durch Beobachtung der Bahnen kann jedoch nicht entschieden werden, ob die Aktivität der Raphe auf der ganzen Länge zwischen den Apices einheitlich ist und ob die Richtung der Aktivität bereits vor dem Kippen bestand oder erst mit dem Kippen die beobachtete Richtung annahm.
Schwenkbewegungen
Recht häufig beobachtet man, dass eine Diatomee, die sich in Valvenlage befindet und vorwärts bewegt, plötzlich stoppt und in eine horizontale Rotation um einen Apex übergeht. Manchmal dreht sie sich dabei um die Apikalachse und liegt danach auf den Gürtelbändern, so dass die Raphen beider Valven die weitere Bewegung bestimmen. Wie oben angemerkt, sollen hier nur die Bewegungsabläufe betrachtet werden, bei denen die Diatomeen dauerhaft auf ihren Valven liegen. Da die Rotation bei dieser Lage typisch aus der Bewegung heraus erfolgt, kann man zwischen einem voreilenden und nachfolgenden Apex unterscheiden. Die Drehung erfolgte bei meinen Beobachtungen immer um den nacheilenden Apex. Da in meinen Kulturen häufig Richtungswechsel erfolgten, so dass die Bahnkrümmung oft nicht feststellbar war, ist ein Zusammenhang von Krümmungsrichtung der Trajektorie vor Beginn der Rotation und Drehrichtung der Diatomee nicht eindeutig nachweisbar. In den gut sichtbaren Fällen erfolgte die Rotation im gleichen Sinne wie die Bahnkurve. Die weitere Bewegung erfolgte, soweit sie lange genug dauerte und nicht durch Stopps unterbrochen wurde, typisch im selben Drehsinn.
 Ich vermute, dass es zur Rotation kommt, wenn der Antriebspunkt nahe dem nacheilenden Apex liegt und die Diatomee mit beiden Raphenästen nahe dem Apex in Kontakt mit dem Substrat kommt. Aufgrund der zwischen den Apices konvexen Valven-Oberfläche liegt dann der voreilende Apex weiter vom Substrat entfernt als der nacheilende Apex (siehe Bild links). Die Apices sind in dieser Lage nicht gleichzeitig fokussierbar.
Ich vermute, dass es zur Rotation kommt, wenn der Antriebspunkt nahe dem nacheilenden Apex liegt und die Diatomee mit beiden Raphenästen nahe dem Apex in Kontakt mit dem Substrat kommt. Aufgrund der zwischen den Apices konvexen Valven-Oberfläche liegt dann der voreilende Apex weiter vom Substrat entfernt als der nacheilende Apex (siehe Bild links). Die Apices sind in dieser Lage nicht gleichzeitig fokussierbar.
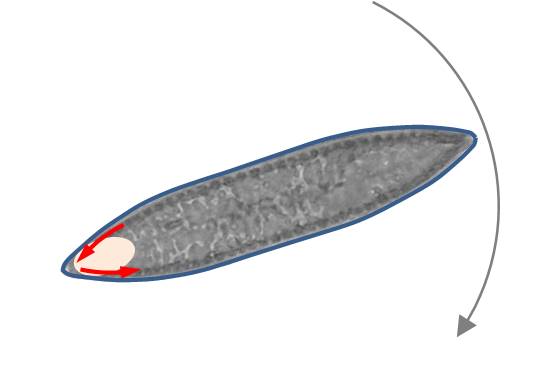 Wie diskutiert wurde, erfolgt die Aktivität der Raphen-Äste meist in entgegengesetzter Richtung, so dass es zur Rotation um die Kontaktfläche kommt. Die Diatomee dreht sich im entgegengesetzten Sinn zum EPS-Transport der Raphen. Dies ist im Bild links skizziert. Dabei wurde die Kontaktfläche durch die ovale Fläche angedeutet. Die blauen kurzen Pfeile zeigen die Transportrichtung der Raphe an.
Wie diskutiert wurde, erfolgt die Aktivität der Raphen-Äste meist in entgegengesetzter Richtung, so dass es zur Rotation um die Kontaktfläche kommt. Die Diatomee dreht sich im entgegengesetzten Sinn zum EPS-Transport der Raphen. Dies ist im Bild links skizziert. Dabei wurde die Kontaktfläche durch die ovale Fläche angedeutet. Die blauen kurzen Pfeile zeigen die Transportrichtung der Raphe an.
Typische Drehungen sind in den nachfolgenden Ausschnittvideos zu sehen. Im Video links unten (150-facher Zeitraffer) durchläuft die Diatomee eine mehrfache sigmoide Kurve, bevor sie in Rotation übergeht. Vermutlich berührt sie abwechselnd mit den gegenüberliegenden Raphen-Hälften das Substrat. Gleich mehrere Rotationen sind im Video rechts daneben (1500-facher Zeitraffer) zu erkennen.
Beobachtung von der Seite
Im Abschnitt über Beobachtung aus horizontaler Blickrichtung wird über eine Methode berichtet, mit der man Diatomeen aus nahezu horizontaler Sicht beobachten kann. Man betrachtet sie so, wie sie ein winziger Beobachter, der auf dem Substrat steht, sehen würde. Bei einem glatten Substrat erkennt man auch das Spiegelbild der Diatomee. Das Video links zeigt in 30-fachem Zeitraffer eine solche Aufnahme von Surirella biseriata. Die Diatomee bewegt sich unter anderem in mehreren S-Kurven. Dabei ändert sich auch die seitliche Neigung der Diatomee. Der Zusammenhang zwischen Neigung und Bahnkrümmung ist in dieser Perspektive allenfalls grob erkennbar.
Es ist mir hoffentlich gelungen, zu zeigen, dass die vorgestellten Analysemethoden ein besseres Verständnis der Trajektorien von Diatomeen ermöglichen. Insbesondere erlauben sie es, das Zusammenspiel zwischen der Aktivität an der Raphe und dem Bewegungsmuster zu untersuchen.
“The Diatoms: Biology and Morphology of the Genera
F. E. Round; R. M. Crawford; D. G. Mann
Cambridge University Press; 1 edition (2007)