
")


Movement patterns of floating Nitzschia sigmoidea
As the videos of the movement illustrate, the motion sequences are fundamentally different from movement on a substrate. On the water surface there is no interaction with a solid substrate, which determines the paths of benthic diatoms. Here, the attractive interaction of the hydrophobic apices also occurs. If there were only these attractive interactions, a distribution on the surface would be similar to that of Lycopodium powder, whereby the movement would come to rest when a local energy minimum was reached. "Local" means that the energy cannot be reduced by a small change. An absolute minimum is not reached because of the stability of the local minima and the active movement of the diatoms. Lattice structures like in crystals do not occur.
The pattern of the diatoms can be described by undirected graphs. The apices form the nodes. Parallel diatoms represent multiple edges (multigraph). If a diatom touches a second diatom at a point other than its apex, there is no bond. According to the simple model of two hydrophobic particles in the distance of the apices one expects structures with few unconnected nodes (i.e. unbound apices). For example, if three diatoms are within the range of the surface forces, the configurations where all ends have bonds to other diatoms should prevail:
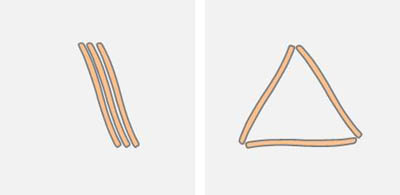
Energetically unfavorable are patterns with free ends, as are shown below:

A chain of three diatoms should close to a triangle because of the attractive interaction of the free ends. The diatoms however, are not passive floating objects, but show active movement. The diatoms release energy and thus temporarily produce patterns with a higher energy.
There are elementary sequences which are observable locally in the entire structure pattern:
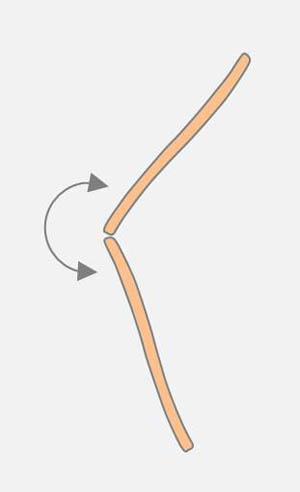
 The end of a diatom can be moved by another diatom along its raphe (see drawing on the left). An existing connection at an apex of the moved diatom can be broken.
The end of a diatom can be moved by another diatom along its raphe (see drawing on the left). An existing connection at an apex of the moved diatom can be broken.
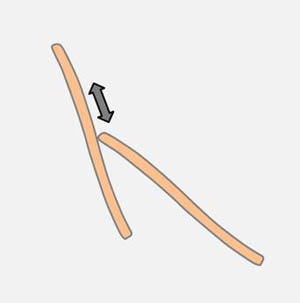
Very frequent are angular changes at connected apices (see drawing on the right). Diatoms that adhere with the apex on a substrate are capable of performing pivoting around the cell pole. It is probably the same mechanism which is observed on the surface of the water.
The static attractive interaction of the apices, the active movement along the raphe and the active change in the angle between two connected diatoms can produce a huge amount of patterns and sequences in a system of several diatoms. In the time lapse video (30x time lapse) at the bottom left we see a cluster of only four diatoms which, by means of the described types of motion, take different forms successively, the number of free ends varying between 2 and 5. To the right of it, another example of motion processes (32x time lapse) is presented, whereby some particles floating on the surface illustrate the activity of the raphes.
Older cultures are often contaminated with EPS. This can also drift on the surface and be transported by the raphes. In the following video on the left (32x time lapse) there are a diatom and two particles. Due to the extruded EPS new patterns of movement on the surface of the water are caused. On the right side, a video with a large time lapse factor (64) shows two diatoms, which have coordinated sequences in their movement.
Although no EPS is recognizable, I suspect EPS as the cause. The two diatoms could be in contact with the lump and transport it along their raphes, which lead to mutual interaction. I would not assume that there is a long-distance effect which is caused e.g. by water vortices.
More difficult is the interpretation in another case. In the video on the left (80x time lapse) you see in phase contrast many bacteria on the surface, which make the water movement visible. The raphe appears to generate a water flow, which is causally connected with the movement. EPS on the raphe and between the diatoms is not recognizable. However, I do not want to rule out this possibility. In this context, it should be mentioned that even individual diatoms are sometimes capable of small movements on the surface of the water. I cannot judge whether lumps of EPS at the Raphe cause this movement.
Light intensity also plays an important role. Thus, in a culture which is exposed to a low intensity of light, parallel diatoms are frequently found. Under the more powerful microscope illumination the activity increases at the raphes and the structure changes into an irregular structure.
Diatoms of diverse localities differ in terms of the movement sequences and the accuracy of the connections at the apices. I suspect a differently strong hydrophobic interaction as reason. Against this background it is worth watching the compilation of the videos.
Due to the mechanical coupling of many diatoms, water flow, activity of the diatoms at their pole and raphe a complex pattern is generated that change in time. I believe that the given presentation is only for discussion. It should not give the impression that all processes of the movement can be explained with the illustrated approaches. Some aspects require further investigation.