
")


Abläufe der Bewegung schwimmender Nitzschia sigmoidea
Wie die Videos der Bewegung illustrieren, unterscheiden sich die Abläufe der Bewegung grundlegend von einer Bewegung auf einem Substrat. Auf der Wasseroberfläche gibt es keine Wechselwirkung mit einem festen Substrat, welche bei benthisch lebenden Diatomeen die Bahnen bestimmen. Hier tritt zusätzlich die attraktive Wechselwirkung der hydrophoben Apizes auf. Gäbe es nur diese attraktive Wechselwirkung, so würde eine Verteilung auf der Oberfläche ähnlich dem von Bärlappsporen entstehen, wobei die Bewegung zur Ruhe käme, wenn ein lokales Energieminimum erreicht ist. „Lokal“ bedeutet, dass die Energie nicht durch eine kleine Veränderung verkleinert werden kann. Ein absolutes Minimum wird wegen der Stabilität der lokalen Minima und der aktiven Bewegung der Diatomeen nicht erreicht. Gitterstrukturen wie bei Kristallen treten nicht auf.
Die Anordnung der Diatomeen kann durch ungerichtete Graphen beschrieben werden. Die Apizes bilden dabei die Knoten. Parallel liegende Diatomeen stellen Mehrfachkanten dar (Multigraph). Berührt eine Diatomee eine zweite Diatomee an einer anderen Stelle als an deren Apex, so sind sie nicht verbunden. Entsprechend dem einfachen Modell von zwei hydrophoben Partikeln im Abstand der Apizes erwartet man Strukturen mit wenigen nicht verbundenen Knoten, also ungebundenen Enden. Befinden sich zum Beispiel drei Diatomeen innerhalb der Reichweite der Oberflächenkräfte, so sollten die Konfigurationen vorherrschen, bei denen alle Enden Bindungen zu anderen Diatomeen aufweisen:
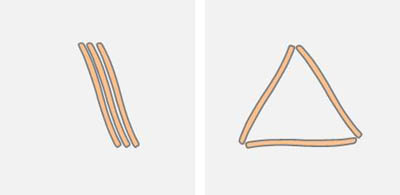
Energetisch ungünstiger sind Anordnungen mit freien Enden, wie sie nachfolgend dargestellt sind:

Eine Kette aus drei Diatomeen sollte sich auf Grund der attraktiven Wechselwirkung der freien Enden zum Dreieck schließen. Die Diatomeen sind jedoch nicht passiv treibende Gebilde, sondern sie zeigen aktive Bewegung. Die Diatomeen geben Energie ab und erzeugen so temporär Muster mit höherer Energie.
Es gibt elementare Abläufe, die lokal im gesamten Gebilde beobachtbar sind:
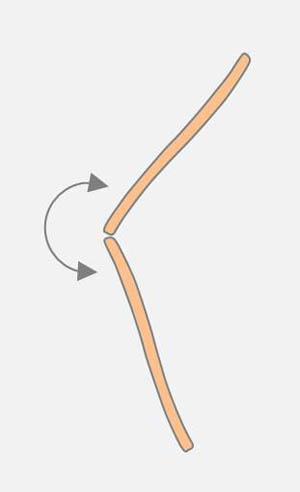
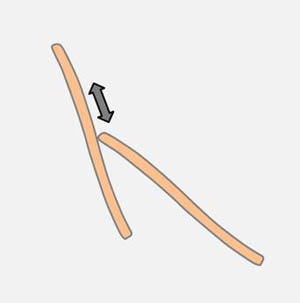 Das Ende einer Diatomee kann von einer anderen Diatomee entlang ihrer Raphe bewegt werden (Zeichnung links). Dabei kann eine bestehende Verbindung an einem Apex der bewegten Diatomee aufgebrochen werden. Sehr häufig sind Winkeländerungen zwischen verbundenen Apizes. Diatomeen, die am Apex an einem Substrat haften, sind in der Lage, Schwenkbewegungen um den Pol der Zelle auszuführen (Zeichnung rechts). Vermutlich handelt es sich um denselben Mechanismus, den man an der Wasseroberfläche beobachtet.
Das Ende einer Diatomee kann von einer anderen Diatomee entlang ihrer Raphe bewegt werden (Zeichnung links). Dabei kann eine bestehende Verbindung an einem Apex der bewegten Diatomee aufgebrochen werden. Sehr häufig sind Winkeländerungen zwischen verbundenen Apizes. Diatomeen, die am Apex an einem Substrat haften, sind in der Lage, Schwenkbewegungen um den Pol der Zelle auszuführen (Zeichnung rechts). Vermutlich handelt es sich um denselben Mechanismus, den man an der Wasseroberfläche beobachtet.
Die statische attraktive Wechselwirkung der Apizes, die aktive Bewegung entlang der Raphe und die aktive Änderung des Winkels zwischen zwei verbundenen Diatomeen kann in einem System mehrerer Diatomeen eine riesige Menge von Mustern und Abläufen erzeugen. Im Zeitraffervideo (30-facher Zeitraffer) unten links sieht man eine Gruppe von vier Diatomeen, die durch die beschriebenen Bewegungstypen nacheinander unterschiedliche Formen annehmen, wobei die Zahl der freien Enden zwischen 2 und 5 schwankt. Rechts daneben ist ein weiteres Beispiel von Bewegungsabläufen (32-facher Zeitraffer) gezeigt, wobei einige an der Oberfläche treibende Partikel die Aktivität der Raphen verdeutlichen.
Ältere Kulturen sind häufig mit EPS verunreinigt. Dieses kann auch an der Oberfläche treiben und an den Raphen transportiert werden. Im nachfolgenden Video links (32-facher Zeitraffer) befinden sich eine Diatomee und zwei Partikel. Durch die EPS-Ausscheidungen treten neue Bewegungsmuster an der Wasseroberfläche auf. Rechts daneben zeigt ein Video in starkem Zeitraffer (64-facher Zeitraffer) zwei Diatomeen, die koordinierte Abläufe in ihrer Bewegung besitzen.
Obwohl man kein EPS erkennt, vermute ich dies als Ursache. Die beiden Diatomeen könnten Kontakt zu dem Klumpen haben und an ihren Raphen transportieren, was zur gegenseitigen Beeinflussung führt. Ich würde nicht von einer Fernwirkung ausgehen, die etwa durch Wasserwirbel verursacht wird.
Schwieriger ist die Interpretation in einem anderen Fall. Im Video links (80-facher Zeitraffer) sieht man im Phasenkontrast viele Bakterien an der Oberfläche, welche die Wasserbewegung sichtbar machen. An den Raphen scheinen Strömungen an der Oberfläche zu entstehen, die ursächlich mit der Bewegung zusammenhängen. EPS an den Raphen und zwischen den Diatomeen ist nicht zu erkennen. Dennoch möchte ich diese Möglichkeit nicht grundsätzlich ausschließen. In diesem Zusammenhang sei erwähnt, dass auch einzelne Diatomeen manchmal zu einer geringen Eigenbewegung an der Wasseroberfläche in der Lage sind. Ob Klumpen von EPS an der Raphe diese Bewegung verursachen, kann ich nicht beurteilen.
Auch die Lichtintensität spielt eine große Rolle. So kann man bei einer Kultur, die einer nur geringen Lichtintensität ausgesetzt ist, häufig parallel verbundene Diatomeen vorfinden. Unter der stärkeren Mikroskop-Beleuchtung steigt die Aktivität an den Raphen und die Struktur geht in eine unregelmäßige Struktur über.
Diatomeen unterschiedlicher Fundorte unterscheiden sich weiterhin hinsichtlich der Bewegungsabläufe und der Exaktheit der Verbindungen an den Apizes. Ich vermute als Grund eine unterschiedlich starke hydrophobe Wechselwirkung. Vor diesem Hintergrund lohnt es, die Zusammenstellung der Videos anzusehen.
Auf Grund der mechanischen Kopplung vieler Diatomeen, Strömungen, der Aktivität der Diatomeen im Bereich der Polporen sowie der Raphen, vorhandenem EPS ergibt sich ein komplexer zeitlicher Verlauf der Muster. Die gegebene Darstellung halte ich nur für eine Diskussionsgrundlage. Es sollte nicht der Eindruck erweckt werden, dass alle Abläufe der Bewegung mit den dargestellten Ansätzen erklärt werden können. Einige Aspekte erfordern weitere Untersuchungen.