
")


 |
|
| Cymbella spec. im Dunkelfeld (40-facher Zeitraffer) | Visualisierung der Bewegung der Diatomeen aus dem Video durch Bildung des Maximums über alle Frames (zum Vergrößern anklicken) |
Bewegung
Etliche benthische Arten verfügen über die Fähigkeit zur Eigenbewegung. Von ihnen besitzen fast alle eine Raphe. Auf glattem Boden gleiten sie in geraden oder gekrümmten Bahnen, wobei die Form der Bahn von der Krümmung der Raphe abhängt. Sie zeigen auch komplexe Bewegungsabläufe wie plötzliche Richtungsumkehr, Drehen um die Apikalachse („Längsachse“), Aufrichten, horizontale Drehbewegung um einen Punkt der Zelle, Pirouetten im aufgerichteten Zustand etc. Insgesamt wirken die Abläufe zufällig und es wird nicht unbedingt ersichtlich, welcher Nutzen daraus hervorgeht.
Als Nutzen kommen insbesondere in Betracht:
- Optimierung der Lichtverhältnisse, denn viele motile Arten zeigen eine positive oder negative Phototaxis. Auch eine Photophobie kann beobachtet werden, bei der Diatomeen auf stark lokale Änderungen der Lichtintensität mit Umkehr der Bewegungsrichtung reagieren.
- Periodische vertikale Wanderung von Diatomeen, die Sandablagerungen insbesondere in Gezeitenzonen bewohnen. Diese Ablagerungen können durch Gezeiten und Strömungen gestört werden. (siehe Übersichtsartikel Harper (1977))
- Aufsuchen von Orten mit besserer Nährstoffkonzentration oder anderen günstigen chemischen Umgebungsverhältnissen (Chemotaxis). In der Veröffentlichung von Karen Grace V. Bondoc et al (2016) wird gezeigt, dass es bei Seminavis robusta eine Bewegung hin zu einer Silikat-Quelle gibt.
- Besiedlung neuer Habitate
- Suche und Annäherung an einen Partner zur geschlechtlichen Fortpflanzung. Die Struktur und Funktion von Sexualpheromonen wurde für Seminavis robusta aufgeklärt (Frenkel, Johannes. PhD Thesis (2014) und Bondoc et al. (2016)).
- Verlassen der Kopulations-Hülle bei bestimmten Spezies (siehe Beitrag zur sexuellen Fortpflanzung).
Eine Annäherung an das Ziel der Bewegung erreichen Diatomeen durch Variation der Bewegungsaktivität, insbesondere durch Steuerung der Dauer der Bewegung in einer Richtung.
An dieser Stelle möchte ich aus meiner Sicht eine Anmerkung machen. Allen genannten Vorteilen der Bewegung ist gemeinsam, dass sie in einer Ortsveränderung liegen. Wenn man die Bewegung mancher Arten betrachtet, so kann man daran zweifeln, ob das immer die Motivation sein kann. Cymatopleura elliptica rotiert meist langsam um eine vertikale Achse, wechselwirkt unkontrolliert mit der Umgebung und kommt dabei kaum vom Fleck (siehe Video). In vielen Fällen ist der Nutzen einer Beweglichkeit nur temporär gegeben wie bei der geschlechtlichen Vermehrung. Dennoch ist die Mehrzahl der Diatomeen in einer Kultur unabhängig von ihrer Größe auf Wanderschaft. Der Nutzen könnte in bestimmten Fällen auch physiologische Hintergründe wie die Regulierung des Energiehaushalts haben.
Seit der Entdeckung der Bewegung von Diatomeen ringt man um ein Verständnis des Bewegungsmechanismus. Ehrenberg beschrieb bereits 1838 eine schneckenartige Sohle zum Kriechen (Die Infusionsthierchen als vollkommene 0rganismen. 1838 p. 175), was sich jedoch nicht als richtig erwies. Bis heute gibt es zahllose Theorien, von denen keine endgültig bestätigt ist. Sie reichen von einer Bewegung durch Kapillareffekt über eine Cytoplasmaströmung bis zum Rückstoßprinzip. Heute werden meist eine Absonderung von Schleim entlang der Raphe und eine Bewegung des Schleims entlang der Raphe als Erklärung herangezogen, wobei Muskelproteine als Antrieb dienen. Die vorherrschende Hypothese stammt von Edgar, L.A. & Pickett-Heaps J.D. (1984). Eine kompakte Darstellung wurde von Menzel, D. and O. Vugrek (1997) erstellt und ist nachfolgend mit der Originalabbildung und originalem Text wiedergegeben:
|
|
| Schematische Darstellung der Gleitbewegung bei pennaten Diatomeen. (a) Unterseite einer wandernden Zelle. Von der hinteren Raphe wird eine Schleimspur gebildet. (b) Querschnitt entlang der Linie in (a). Die verkieselte Zellwand besteht aus zwei Teilen (blaugrau), welche wie die beiden Hälften einer Petrischale übereinandergestülpt sind. Die Chloroplasten sind grün dargestellt, der Kern in der Mitte der Zelle ist der Übersichtlichkeit halber weggelassen. Unterhalb der Raphen befindet sich jeweils ein Paar von Aktinfilamentbündeln (blau), die als Bahnen für den Transport von Schleimvesikeln (rosa) dienen. Die Vesikel verschmelzen an den Enden der Raphen mit der Plasmamembran. Jedes Vesikel entlädt einen Schleimfaden (rot) an die Außenseite der Membran, wo er aufquillt und durch den Spalt der Raphe gedrückt wird. Ein Ende des Fadens haftet am Untergrund fest, das andere bleibt an die Zellmembran gebunden, und man nimmt an, daß dieses Ende mit einem Motormolekül (dunkelviolett) verbunden ist, das den Transport des Fadens entlang der Aktinbündel übernimmt. Obere und untere Raphe produzieren gleichzeitig Schleimfäden. (c) Ausschnitt aus b. |
| Menzel, D. and O. Vugrek, Muskelproteine in Pflanzenzellen. Biologie in unserer Zeit, 1997. 27(3): p. 195-203. Copyright Wiley-VCH Verlag GmbH & Co. KGaA. Reproduced with permission. |
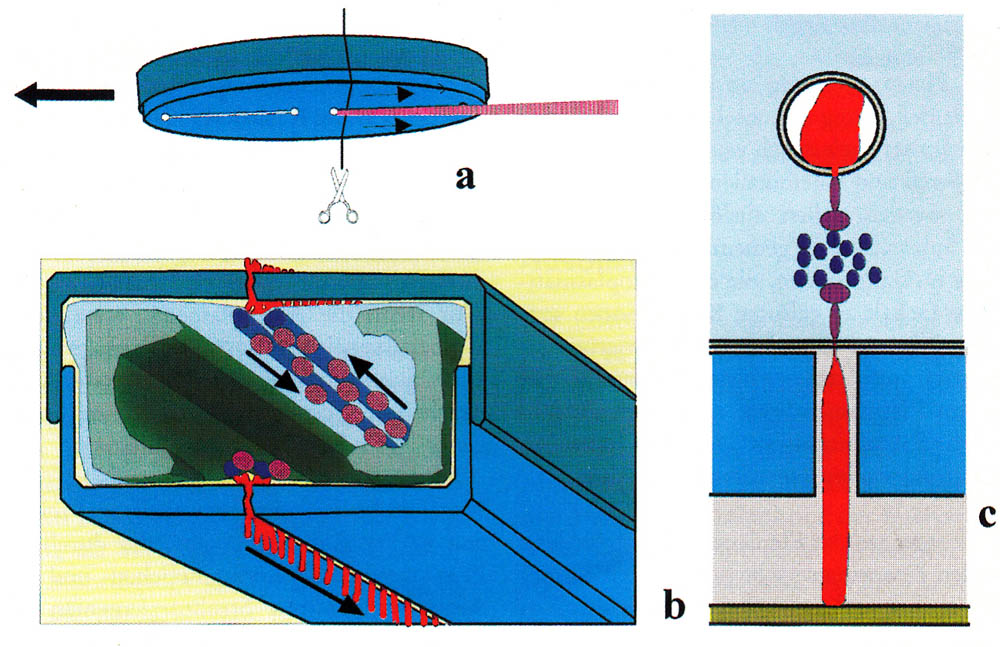
Zwei bekannte Probleme dieser Theorie seien an dieser Stelle erwähnt:
- Die Bewegung von Myosin auf Aktinfilamenten erfolgt stets in einer Richtung. Die Bewegung der Diatomeen wechselt jedoch die Bewegungsrichtung, wobei es keinen Unterschied in der Geschwindigkeit gibt. Diatomeen haben ein oder zwei Raphensysteme auf einer Valve. Jedes Raphensystem erlaubt Bewegungen in beiden Richtungen. Bei manchen Spezies kann man sogar beobachten, wie Partikel in entgegengesetzter Richtung entlang derselben Raphe transportiert werden und kollidieren (Nultsch, W. (1962)).
- Bei vielen motilen Diatomeen ist die Raphe nicht einfach ein offener Schlitz. Manche bewegen sich trotz einer nahezu geschlossenen Nut-und Feder-Raphe. Andere bewegliche Spezies schaffen das mit einer Kanalraphe, die auf den Valven aufsitzt und nur durch eine Serie von Poren mit dem Inneren der Schale verbunden ist.
Zumindest sind Erweiterungen erforderlich, um die Beobachtungen durch den beschriebenen Actin-Myosin-Transport zu erklären.
J. Wang, S. Cao, C. Du und D. Chen haben die Bewegung bei Navicula sp. untersucht und schlagen ein aktualisiertes Modell vor. In diesem Modell wird die Diatomee durch Scheinfüßchen bewegt, die aus der Valve ragen. Man kann nicht zwangsläufig davon ausgehen, dass dieses Modell auf andere Gattungen übertragbar ist.
Auf dieser Homepage werden zu den bereits existierenden zahllosen Beobachtungen zum Bewegungsverhalten von Diatomeen nach und nach weitere hinzugefügt. Entsprechend meiner Möglichkeiten sind dies stets Beobachtungen mit dem Lichtmikroskop und man kann sich fragen, was man nach etwa 200 Jahren lichtmikroskopischer Untersuchung überhaupt noch beobachten will. Tatsächlich wird vieles nachvollzogen, anderes regt den Leser vielleicht zu eigenen Beobachtungen an. Im Einzelfall konnte ich nicht prüfen, wie weit diese in der Literatur beschrieben sind. Für diesbezügliche Hinweise wäre ich dankbar. Solche Beobachtungen können keine neue Erklärung des Mechanismus der Bewegung liefern, aber möglicherweise einen kritischen Blick auf die beschriebene Modellvorstellung erlauben.
Edgar, L.A. & Pickett-Heaps J.D. (1984), Diatom locomotion., Progress in Phycological Research Vol. 3: 47-88
Frenkel, Johannes. PhD Thesis (2014). Struktur und Funktion von Sexualpheromonen der Diatomee Seminavis robusta. Friedrich-Schiller-Universität Jena, Biologisch-Pharmazeutische Fakultät
Harper, M.A. (1977). Movements. In: The Biology of Diatoms, (D. Werner, ed). 224-249, Blackwell, Oxford
Karen Grace V. Bondoc, Jan Heuschele, Jeroen Gillard, Wim Vyverman & Georg Pohnert. Selective silicate-directed motility in diatoms. Nature Communications http://dx.doi.org/10.1038/ncomms10540
Bondoc, Karen Grace & Lembke, Christine & Vyverman, Wim & Pohnert, Georg. (2016). Searching for a Mate: Pheromone-Directed Movement of the Benthic Diatom Seminavis robusta. Microbial Ecology. 72. 10.1007/s00248-016-0796-7.
Siehe auch:
Jeroen Gillard, Johannes Frenkel, Valerie Devos, Koen Sabbe, Carsten Paul, Martin Rempt, Dirk Inzé, Georg Pohnert, Marnik Vuylsteke, Wim Vyverman: Metabolomik unterstützt die Strukturaufklärung eines Sexualpheromons von Kieselalgen., Angewandte Chemie, DOI: 10.1002/ange.201208175
Nultsch, W. (1962) Über das Bewegungsverhalten der Diatomeen., Planta 58: 22.
Wang, J., Cao, S., Du, C. & Chen, D. Underwater locomotion strategy by a benthic penate diatom Navicula sp. Protoplasm 250, 1203–1212 (2013).