
")


| Surirella biseriata (600x and 700x time lapse) | Almost circular path (150x time lapse) |
Curvature of the trajectories by the example of Surirella biseriata
In April 2017 I isolated a diatom of the genus Surirella in the creek Lein (48°53'04.6"N 9°38'29.2"E), the outflow from the Aichstruter reservoir.
 |
The picture on the left shows the diatom in valve view. I assign them to the species Surirella biseriata (see http://cyclot.sakura.ne.jp/keisougazou/surirel/Surirell/sursei.html). The video above left shows an example of the movement of diatoms in a culture in 600x time-lapse. These diatoms usually move very slowly. In Petri dishes made of polystyrene, the fastest movements were around 2.8 µm per second. In order to cover their own length, which in my cultures was typically a bit over 210 µm, they needed more than one minute. Observations on vertical glass surfaces showed significantly higher velocities.
As the video at the top left shows, those Surirella biseriata often only move a little bit back and forth. Even in moderately densely populated cultures, a noticeable EPS film is formed on the substrate. Diatoms glide on it at the slightest movement of the petri dishes and their active mobility is limited. To observe the movement, it is therefore advisable to pipette a sufficient number of diatoms into a fresh petri dish and observe them there. In my opinion, longer pathways are also covered mainly by diatoms, which have not yet stored many reserve materials. For longer sections of the path without reversal of direction, a high light intensity does not seem to be favourable.
If the diatoms do not travel very short distances between reversal points, it is obvious that the trajectory is curved. Between the reversal points there is clearly always one of the raphes in contact with the flat substrate. As the curvature of the raphe is uniform on each side of the diatom, the curvature of the trajectory can undoubtedly be assigned to one of the two raphe sections between the apices. At the top right you can see an example where the diatom moves approximately on a circular path. For purposes of clarity, a circle is shown. The video ends with a superposition of all frames. Apparently, the curvature of the path is determined by the exterior part of the raphe.
Surirella biseriata can be found in valve view (valve is uppermost) and girdle view (girdle is uppermost). When the diatoms rotate around the apical axis, which is often observed, they change frequently between these positions. Collisions between diatoms can also cause a change of position. Only the movement in valve view is to be considered, as here well observable and describable paths are run through. Particularly remarkable are the curvatures at the reversal points.
The prerequisites for an analysis of the tracks as presented in the post "Analysis of Trajectories I" are not given due to the strongly curved raphes on the outer edge. However, this geometry suggests a simple interpretation of movement patterns.
Reversal without changing the direction of curvature
In this case, a diatom, which describes a path in a clockwise direction by viewing from above, continues to move in a clockwise direction after reversing. The same applies to the counterclockwise direction.
In the sketch below on the left side two such reversal points are shown in sequence. The raphe which is active in the sections is marked in red. It is in mechanical contact with the substrate.
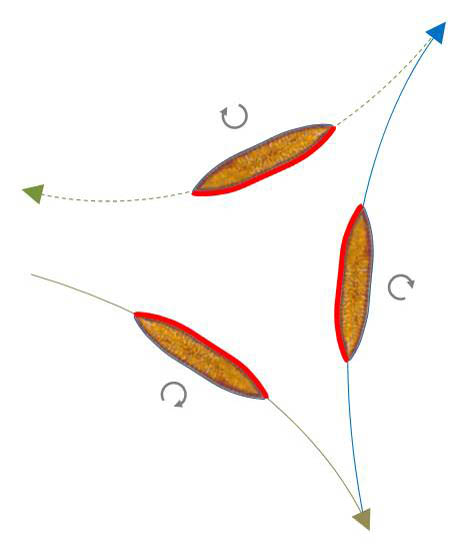 |
From the trajectory nothing can be stated about the activity of the other section of the raphe-system. Obviously the side that is in contact with the substrate changes when reversing (tilting around the apical axis). In addition, the direction of the activity of the raphe sections is different, resulting in a reversal of movement. To the right of it is a 300x time-lapse video showing such a reversal.
 |
At the end of the short video, an image is displayed that shows the whole trajectory and that was created by superimposing all frames of the video.
This form of reversal with keeping the turning sense is very common. The picture on the left (click to enlarge) shows a superimposition of the frames of a video of the movement in a culture. In many places you can see the characteristic patterns of this behaviour.
It follows from this that very often the movement in the raphe halves is contrary. The activity of the entire raphe system is carried out in one direction of rotation, so to speak in a "circle".
Reversal with changing the direction of curvature
This form of reversal was presented as a typical form in Navicula. In Surirella biseriata, it is sufficient for the direction of the raphe's activity to reverse in case of continuous contact between the raphe and the substrate. The sketch on the left below illustrates the process.
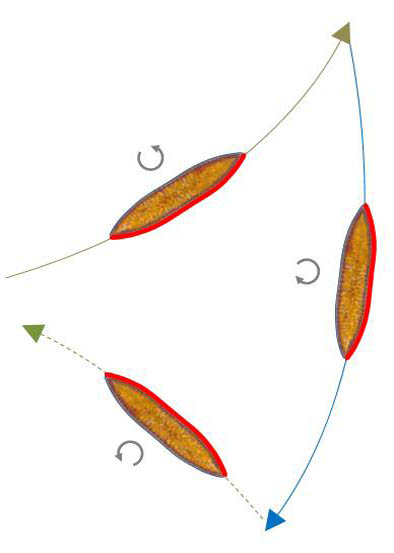 |
In the video to the right (150x time lapse) you can see a reversal without changing the direction of rotation, as already introduced. Then the direction of the activity is changed. At the end of the video, the superimposition of frames is displayed again. I could observe this kind of change of direction only occasionally. This may also be due to the fact that such changes of direction occur in short succession, so that the curvature of the path between the reversal points is not recognizable.
Sigmoid trajectories
Round et. al (2007) are stating (p. 105): „Movement is directional, the path taken correspondingly fairly closely to the course of the raphe system (..) – curved where the raphe is curved (e.g. some Nitzschia species with eccentric raphe systems), straight where the raphe is straight (e.g. Navicula, Pinnularia), and even sigmoid where the raphe is sigmoid (e.g. Pleurosigma angulatum).”
Pleurosigma and Gyrosigma have a raphe-system on each valve, which represents a sigmoid structure with the inflection point in the middle of the valve. A sigmoid path is run through when the centre of force changes from a section with a positive curvature to one with a negative curvature and the direction of travel is maintained. The same applies to the reverse change from negative to positive curvature. Apparently, it is not the position of the raphe-branches that is important but the existence of positive and negative curvatures. A sigmoid raphe-system is therefore not necessarily required. Therefore, the occurrence of sigmoid trajectories can be assumed in Surirella biseriata. A change of the mechanical contact to the substratum from one side to the other is sufficient, as long as the two raphe-branches work in the same direction as the sketch below left shows. The trajectory therefore has a inflexion point at the location of the change of contact.
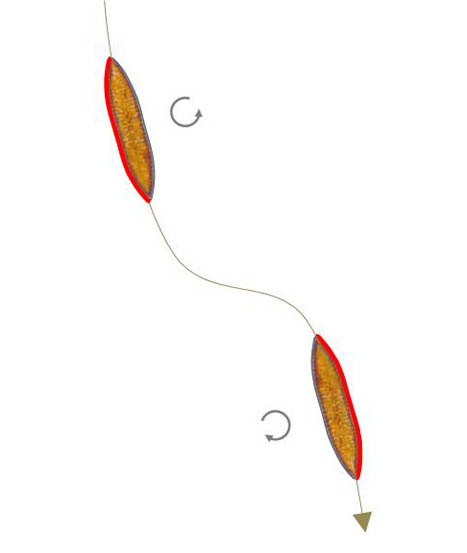 |
In the video next to it (900x time lapse) such a path followed by a superposition of all frames can be seen.
The reversal of the direction of movement described above without changing the direction of curvature of the trajectory (maintaining the sense of rotation) and the occurrence of inflection points are based on the change of the mechanical contact between the substrate from one half of the raphe to the other. Reversal with keeping the direction of rotation is much more frequent than the occurrence of inflection points. One can conclude from this that the activity of the raphe-segments on a valve after tilting is usually opposite and not parallel. By observing the trajectories, however, it is not possible to determine whether the activity of the raphe is uniform along the entire length between the apices and whether the direction of the activity already existed before tilting or whether it took the observed direction with tilting.
Pivoting movements
It is often observed that a diatom, which is lying on a valve and moves forward, suddenly stops and starts rotating horizontally around an apex. Sometimes it rotates around the apical axis and then lies on the girdle bands, so that the raphes of both valves determine the further movement. As mentioned above, only the motion sequences in which the diatoms lie permanently on their valves are to be considered here. As the rotation in this position typically takes place out of the movement, one can distinguish between a leading apex and a trailing apex. In my observations, the rotation always took place around the trailing apex. Since in my cultures there were frequent reversals of direction, so that the path curvature was often not noticeable, a correlation between the direction of curvature of the trajectory before the start of rotation and the direction of rotation of the diatom cannot be clearly demonstrated. In the clearly visible cases, the rotation took place in the same sense as the trajectory. Provided that the further movement lasted long enough and was not interrupted by stops, it typically took place in the same direction of rotation.
 I suspect that rotation occurs when the centre of force is close to the trailing apex and the diatom on both rape branches near the apex comes into contact with the substrate. Due to the convex surface of the valves, the leading apex is more distant from the substrate than the trailing apex (see picture on the left). The apices cannot be focused at the same time in this situation.
I suspect that rotation occurs when the centre of force is close to the trailing apex and the diatom on both rape branches near the apex comes into contact with the substrate. Due to the convex surface of the valves, the leading apex is more distant from the substrate than the trailing apex (see picture on the left). The apices cannot be focused at the same time in this situation.
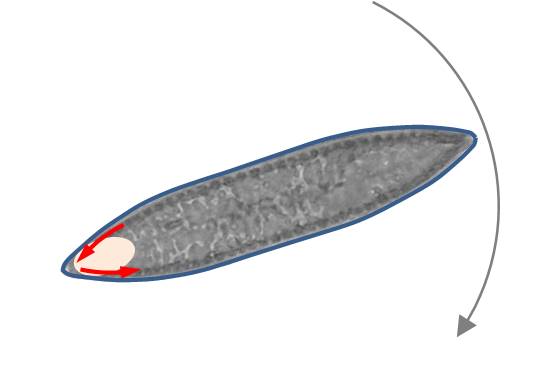 As discussed, the activity of the raphe-branches is usually in the opposite direction, so that it comes to a rotation around the contact surface. The diatom rotates in the opposite direction to the EPS transport of the raphes. This is sketched in the picture on the left. The contact surface was indicated by the oval area. The blue short arrows indicate the direction of transport of the raphe.
As discussed, the activity of the raphe-branches is usually in the opposite direction, so that it comes to a rotation around the contact surface. The diatom rotates in the opposite direction to the EPS transport of the raphes. This is sketched in the picture on the left. The contact surface was indicated by the oval area. The blue short arrows indicate the direction of transport of the raphe.
Typical rotations can be seen in the following videos. In the video at the bottom left (150x time lapse), the diatom runs through a repeated sigmoid curve before it starts rotating. It probably touches the substrate alternately with the opposite raphe halves. Several rotations can be seen in the video to the right of it (1500x time lapse).
Side view observation
In the section on observation with horizontal viewing direction, a method is described that allows diatoms to be observed from a nearly horizontal viewpoint. One looks at them like a tiny observer standing on the substrate would see them. On a smooth substrate you can also see the mirror image of the diatom. The 30x time-lapse video on the left shows such a recording of Surirella biseriata. The diatom moves partly in several S-curves. The lateral inclination of the diatom also changes. The connection between inclination and curvature of path is however not clearly recognizable from this perspective.
I hope to have succeeded in showing that the analytical methods presented allow a better understanding of the trajectories of diatoms. In particular, they allow investigating the interaction between the activity of the raphe and the pattern of movement.
“The Diatoms: Biology and Morphology of the Genera
F. E. Round; R. M. Crawford; D. G. Mann
Cambridge University Press; 1 edition (2007)