")


Lieber Besucher,
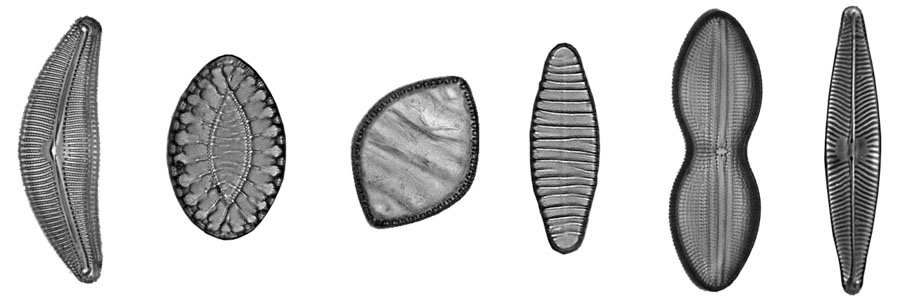
möglicherweise haben Sie schon Bilder von Diatomeen mit ihren vielfältigen Formen und feinen Strukturen gesehen. Vielleicht hatten Sie auch Gelegenheit, lebende Diatomeen unter dem Mikroskop zu beobachten und bemerkt, dass manche in der Lage sind, sich gleitend oder ruckelnd fortzubewegen. Dabei wechseln sie immer wieder die Bewegungsrichtung. Der Mechanismus der Bewegung ist nicht zu erkennen.
In meiner Freizeit beobachte ich Diatomeen mit dem Fokus auf Spezies, die Eigenbewegung zeigen. Mich fasziniert die Reaktion von Diatomeen auf Umweltbedingungen und äußere Reize, wie Hindernisse oder Licht. In Diatomeen-Kulturen versuche ich, den Zusammenhang zwischen Bewegung und der Bildung von Kolonien zu verstehen.
Ich möchte erwähnen, dass ich (Thomas Harbich, siehe Kontakt) Physik studiert habe und mein Arbeitsleben mit typischen Tätigkeiten eines Ingenieurs im Bereich der Telekommunikation und des Automobilbaus verbracht habe. Seit einigen Jahren bin ich im Ruhestand.
Zweck dieser Homepage ist es, zu eigenen Beobachtungen an Diatomeen anzuregen. Falls Sie gerne mikroskopieren und diese Anregung aufgreifen, finden Sie hoffentlich nützliche Hinweise auf diesen Seiten.
Sie erfahren hier etwas über die nachfolgenden Aspekte:
- Kultivierung von Diatomeen
- Beobachtung und Videoaufnahme der Bewegung von Diatomeen in unterschiedlichen Umgebungen
- Auswertung der entstandenen Bilder und Videos
- Hypothesen und Schussfolgerungen aus den Beobachtungen
Da es zur Biologie der Diatomeen eine reichhaltige Literatur und viele Webseiten gibt, beschränke ich mich auf einige Anmerkungen, Links und Literaturhinweise.
Von Zeit zu Zeit soll die Seite um Beobachtungen und Hypothesen ergänzt werden. Wenn sie an dem Thema interessiert sind, dann besuchen sie bitte diese Homepage von Zeit zu Zeit. Ich bin dankbar für inhaltliche Kritik und Verbesserungsvorschläge.
Beachten Sie bitte, dass die Inhalte urheberrechtlich geschützt sind. Wenn Sie etwas von dieser Seite zitieren, so nennen Sie bitte meinen Namen und fügen Sie einen Link auf diese Seiten bei. Bitte sprechen Sie mich an, wenn Sie Bilder oder Videos verwenden wollen. Kommerzielle Interessen verfolge ich nicht.
Die Seite enthält viele kurze Videos. Zu ihrer Betrachtung ist ein Browser erforderlich, der HTML5 beherrscht. Sollte ein Video auf einem Browser nicht korrekt laufen, empfiehlt es sich, einen anderen Browser zu versuchen.
Zur Einstimmung diene die Darstellung von Diatomeen aus dem Werk "Kunstformen der Natur" von Ernst Heinrich Philipp August Haeckel (16.02.1834 bis 09.08.1919):
 |
 |
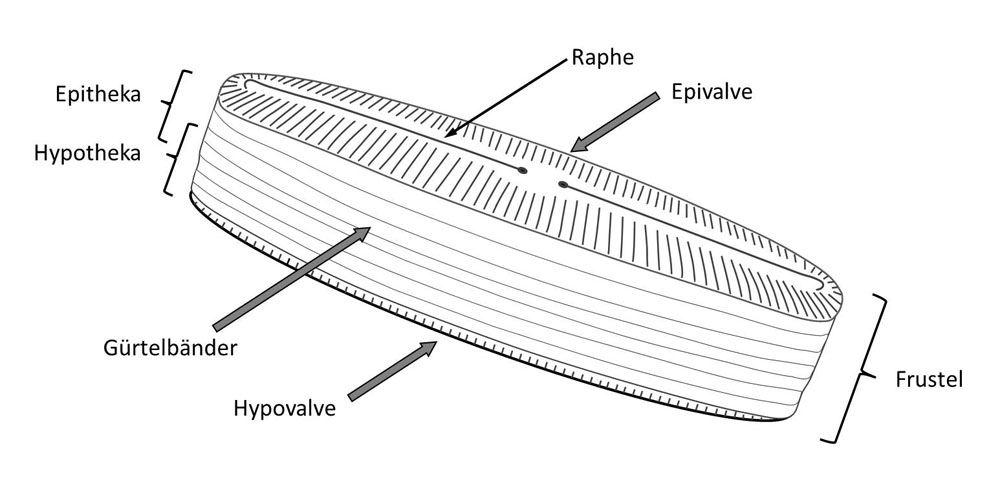
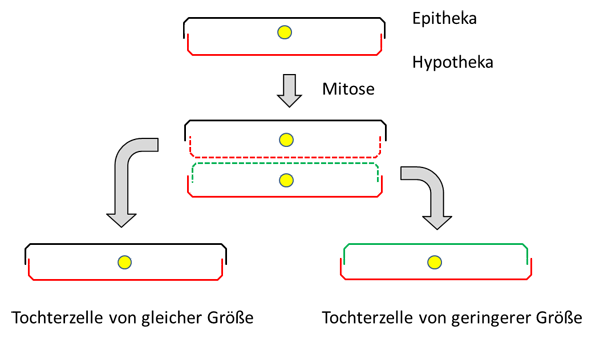

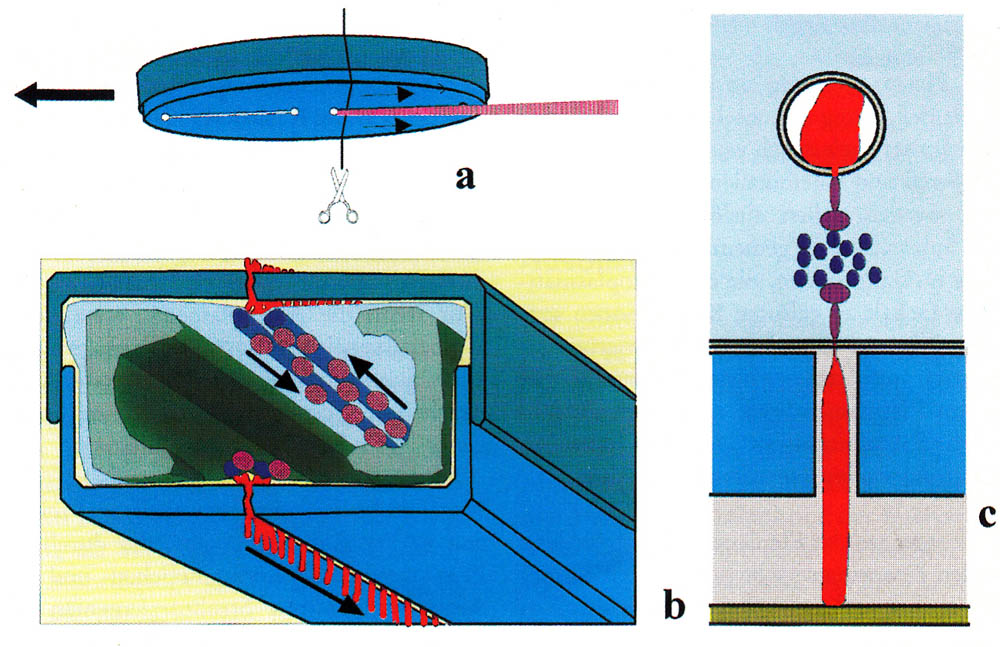
 and Applications) Vadim V. Annenkov (Editor), Richard Gordon (Series Editor), Joseph Seckbach (Series Editor), Wiley-Scrivener; First published: 29 October 2021,
and Applications) Vadim V. Annenkov (Editor), Richard Gordon (Series Editor), Joseph Seckbach (Series Editor), Wiley-Scrivener; First published: 29 October 2021,